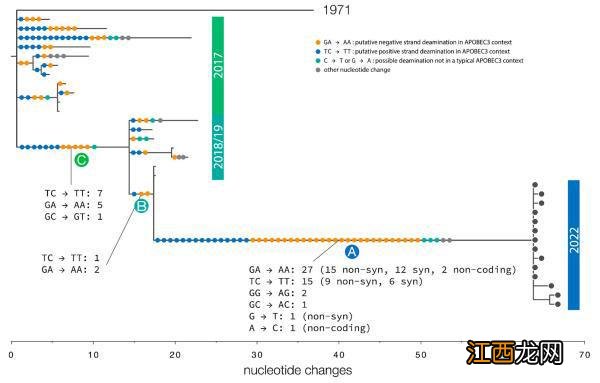
研究者拥有的猴痘病毒的基因组测序样本,包括存档样本和来自非人类动物的感染样本 , 通过对样本的比较,研究者确定了三个不同的进化树分支 。2022年的人类猴痘病毒属于西非分支的一部分,在这一分支中,最古老的基因组来自1971年的尼日利亚,其余的来自2017年以来的采样 。对这些近期基因组的进化树分析也显示了GA→AA和TC→TT突变的倾向,这表明APOBEC3蛋白酶编辑是这组猴痘病毒中观察到的单核苷酸遗传变异的主要来源 。
文章插图
图一:从2017-2022年的人类猴痘感染者采样的猴痘基因组进化树,以及1971年尼日利亚暴发的外群序列 。每个分支上的SNP都用圆圈表示,圆圈的颜色由APOBEC3编辑的TC→TT(蓝色)和GA→AA(橙色),是否为非目标C→T或G→a突变(蓝绿色),或其他SNP类型(灰色)着色 。
【症状轻或致猴痘疫情被低估,病毒隐秘人际传播可能已超五年】
研究者在这篇文章中重点回答了三个关键问题 。首先,在出现在人类有限的传播链中之前,这种假定的APOBEC3编辑是否发生在非人类动物宿主中?
有研究表明,在啮齿类动物中发现的单一APOBEC3蛋白酶表现出对5'TYC的偏好,例如,人类APOBEC3蛋白酶会优先突变CCC > CCT中的C,而小鼠APOBEC3蛋白酶优先发生突变TYC > TYT中的C 。但兰博特等人发现 , 在整个猴痘病毒进化树中142个APOBEC3取代假设中,只有46个出现了这种情况 。
论文进一步表示, 如果基因进化树上显示APOBEC3蛋白酶编辑是人类 , 而不是非人类动物编辑复制的特征 , 那么预计在2017年疫情暴发前将看不到它存在的证据 。为了验证这一点,研究者选择了1970年利比里亚的一个较远的外系病毒分支样本,将2017年猴痘病毒基因组与其比较 , 发现了28个突变 。这个分支上的突变没有表现出如此强的偏向信号,28个单核苷酸多样性位点中只有10个匹配APOBEC3编辑的特征 。因此 , 研究者认为,目前研究者看到的病毒基因组在2017年以后的变异模式是其在人体内进行复制的暗示 。这些变化发生在2017年和2018年之间 , 此后又出现在2022年的病毒样本中,这意味着病毒至少在2017年以来有着持续的人际传播 。符合APOBEC3特征的10个突变可能代表了2017年病例之前的一段额外的人际传播时期 。考虑到该分支的APOBEC3型突变比2017年至2018年期间的APOBEC3型突变要少,这不太可能代表一个很长的时期 。
其次 , APOBEC3蛋白酶编辑机制是否推动了人类猴痘病毒的进化?
APOBEC3蛋白酶的正常作用是防御病毒 。它能够在病毒整个基因组的任意位置诱导单核苷酸出现G→A、C→T的突变,这可能使病毒产生足够的有害变化从而失活 。
在双链DNA病毒中,APOBEC3蛋白酶在病毒基因组被复制和单链暴露时发挥作用 。在病毒重复的复制过程中,任意一条链都可以打开,被APOBEC3蛋白酶脱氨 , 导致正链上C→T和G→A的变化 。因此,被APOBEC3蛋白酶广泛突变的病毒基因组很可能无法存活,也不会进一步传播 。但偶尔,一个被APOBEC3蛋白酶编辑的基因组可能仍然存活并被传播 。猴痘病毒能够在细胞质中进行的大量基因组复制可能意味着其大多数基因组不受APOBEC3编辑作用的影响 。
此外,研究者提出了一个假设,即目前观察到的APOBEC3突变,是自然选择消除了那些会让病毒付出重大适应度代价的突变、残留下来的对病毒危害最小的突变 。
对此,他们考虑了2018年基因组蛋白质编码区域的所有GA和TC二核苷酸位点,这些位点可能是APOBEC3编辑的目标,进而研究了这些位点的突变会产生什么影响 。在21,230个这样的二核苷酸位点中,69.3%(14,707)会发生氨基酸替换,24.7%(5253)会生成同义氨基酸,6.0%(1270)会产生停止密码子 。对于2022年的基因组,在这些二核苷酸发生的40个突变中 , 60%(24)会产生氨基酸替换,40%(16)是同义突变(另外2个APOBEC3突变位于非编码区域) 。在期望值为0.247的二项分布下,研究者得到16个或更少的同义突变的概率为P=0.024 。这在一定程度上证明了研究者的假设 。
相关经验推荐













- 农民在秋季或冬季还有哪些农事活动 农民冬季的农事活动是什么
- 11月15日0-24时眉山新增本土确诊病例1例及无症状感染者1例
- 轻泥土手工制作教程视频 轻泥土手工制作教程
- 11月15日呼和浩特新增本土确诊病例73例、无症状感染者1057例
- 附活动轨迹 11月16日威海环翠区新增1例无症状感染者
- 首期个人养老金可投资基金名录或将发布
- 11月15日0-24时凉山州甘洛县新增省内无症状感染者4例
- 玉米粗缩病用什么药,附症状 什么药治玉米粗缩病
- 猪笼草捕食昆虫的过程 猪笼草捕食昆虫的过程或方法
- 11月16日0-12时珠海新增本土新冠病毒无症状感染者5例